2. Probiotics in the Prevention and Treatment of ADs
Probiotics are administered to treat and prevent various diseases. The probiotics are formulated either in single strain or in combination of strains. Lactobacilli and bifidobacteria are among the commonly applied probiotic LAB used to restore dysbiosis, and to combat infections, allergies, metabolic disorders and autoimmune disorders
| [8] | Al-Salami H, Caccetta R, Golocorbin-Kon S, Mikov M. Probiotics Applications in Autoimmune Diseases. In: Rigobelo E, editor. Probiotics, InTech; 2012. https://doi.org/10.5772/50463 |
[8]
.
Autoimmune disorders occur when the immune system erroneously act against its own body cell by producing antibodies. Consequently, the condition is complicated by inflammation as a result of intensive attack of antibodies on targeted tissue and dramatic damage to tissue, leading to malfunctioning of vital organs. Autoimmune disorders include type 1 diabetes, rheumatoid arthritis, Graves' disease, systemic lupus erythematosus and inflammatory bowel disease (IBD)
| [8] | Al-Salami H, Caccetta R, Golocorbin-Kon S, Mikov M. Probiotics Applications in Autoimmune Diseases. In: Rigobelo E, editor. Probiotics, InTech; 2012. https://doi.org/10.5772/50463 |
[8]
.
According to a previous report
, the incidence of autoimmune disease is increasing by 19.1% annually worldwide. The root causes of autoimmune disease have not been well elucidated; however, genetic (30%) and environmental (70%) predispositions are anticipated to be the main contributing factors for the rapid increase in ADs prevalence
| [8] | Al-Salami H, Caccetta R, Golocorbin-Kon S, Mikov M. Probiotics Applications in Autoimmune Diseases. In: Rigobelo E, editor. Probiotics, InTech; 2012. https://doi.org/10.5772/50463 |
[8]
.
Since ancient times probiotics have been applied to amend gastrointestinal symptoms. The in vivo and invitro studies have shown positive impact of probiotics on the immune system. This accounts for the relationship between immune system and gut microbiome, their polysaccharide antigens, and key metabolites produced by probiotic bacteria. Modification of gut microbiome by probiotics in many randomized control design studies in type 1 diabetes (T1D), rheumatoid arthritis (RA), Graves' disease (GD), systemic lupus erythematosus (SLE) and inflammatory bowel disease (IBD) in animal models and clinical trials is promising alternative to ADs treatment and prevention
.
2.1. Type 1 Diabetes (T1D)
Type 1 diabetes is a chronic autoimmune disorder that occurs when insulin-producing pancreatic β-cells are diminished by excessive antibodies secreted by the immune system and is commonly detected in early childhood. Patients with T1D are largely dependent on exogenously supplied insulin to maintain blood glucose levels. According to research findings, the progression of T1D is exacerbated by the interaction of predisposing HLA genes accompanied by environmental factors, viral infection and intestinal dysbiosis. The prevalence of T1D is alarmingly increasing worldwide at 3–5% annually and is expected to double every 20 years due to inadequate preventive and therapeutic approaches
| [11] | Hara N, Alkanani AK, Ir D, Robertson CE, Wagner BD, Frank DN, et al. The role of the intestinal microbiota in type 1 diabetes. Clinical Immunology 2013; 146: 112–9. https://doi.org/10.1016/j.clim.2012.12.001 |
[11]
.
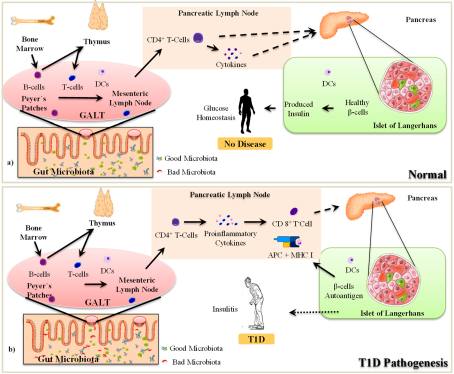
In the pathogenesis of T1D, pancreatic β-cells are destroyed by host immune cells derived from cytotoxic T cell. Regardless of the unknown mechanism by which cytotoxic T cells are activated as autoreactive islets, they cause significant damage to pancreatic β-cells. Following the formation of islets, other immune cells, such as macrophages, access Langerhans islets, leading to inflammation and insulitis (
Figure 1). Consequently, insulitis causes significant damage to β-cells and impaired insulin production
| [12] | Mishra, Wang S, Nagpal R, Miller B, Singh R, Taraphder S, et al. Probiotics and Prebiotics for the Amelioration of Type 1 Diabetes: Present and Future Perspectives. Microorganisms 2019; 7: 67. https://doi.org/10.3390/microorganisms7030067 |
[12]
.
The autoreactive immune response emanates from the pancreatic lymph node, which houses autoreactive T cells and β-cell antigens. Innate immune component macrophages are responsible for the activation of autoreactive cytotoxic T cells. Dendritic cells and macrophages are innate immune effectors that trigger autoreactive cytotoxic T cells. In addition, dendritic cells (DCs) and macrophages possess histocompatibility complex class II (MHC II) molecules to act as professional antigen-presenting cells in activating T cells as part of adaptive immunity in T1D pathogenesis
| [13] | Drexhage HA, Dik WA, Leenen PJM, Versnel MA. The Immune Pathogenesis of Type 1 Diabetes: Not Only Thinking Outside the Cell but Also Outside the Islet and Out of the Box. Diabetes 2016; 65: 2130–3. https://doi.org/10.2337/dbi16-0030 |
| [14] | Paschou SA, Papadopoulou-Marketou N, Chrousos GP, Kanaka-Gantenbein C. On type 1 diabetes mellitus pathogenesis. Endocrine Connections 2018; 7: R38–46. https://doi.org/10.1530/EC-17-0347 |
[13, 14]
. The connection between innate and adaptive immunity implies the predominant role of environmental factors in triggering autoimmune T1D in the presence of genetic predisposition
| [12] | Mishra, Wang S, Nagpal R, Miller B, Singh R, Taraphder S, et al. Probiotics and Prebiotics for the Amelioration of Type 1 Diabetes: Present and Future Perspectives. Microorganisms 2019; 7: 67. https://doi.org/10.3390/microorganisms7030067 |
[12]
.
The whole sum of microbes and their genome in the gastrointestinal tract constitutes the gut microbiota. The diversity of the microbiome is largely dependent on environmental factors such as diet and antibiotic exposure
| [15] | Dovi KS, Bajinka O, Conteh I. Evidence and possible mechanisms of probiotics in the management of type 1 diabetes mellitus. J Diabetes Metab Disord 2022; 21: 1081–94. https://doi.org/10.1007/s40200-022-01006-2 |
[15]
. The consumption of probiotics, for example either in diet or in capsule form, can improve the composition of the microbiota or provide substantial health benefits to the host
| [16] | Zhang X, Zhang Y, Luo L, Le Y, Li Y, Yuan F, et al. The Beneficial Effects of a Multispecies Probiotic Supplement on Glycaemic Control and Metabolic Profile in Adults with Type 1 Diabetes: A Randomised, Double-Blinded, Placebo-Controlled Pilot-Study. DMSO 2023; Volume 16: 829–40. https://doi.org/10.2147/DMSO.S400119 |
[16]
. The valuable aspects of the microbiota include fermentation of carbohydrates, the production of vitamins, the development of gut-associated lymphoid tissues (GALTs), the polarization of gut-specific immune responses and the prevention of colonization by pathogens. The gut microbiome-host interaction plays an important role in the establishment, maturation and regulation of the immune system. It is, therefore, obvious that impaired microbiome composition, so called dysbiosis, and interactions with the host have significant impacts on human health
| [17] | Kamada N, Seo S-U, Chen GY, Núñez G. Role of the gut microbiota in immunity and inflammatory disease. Nat Rev Immunol 2013; 13: 321–35. https://doi.org/10.1038/nri3430 |
[17]
. In the case of T1D, several studies have reported that the gut microbiome and its metabolites are linked to the pathophysiology of T1D before the progression of islet autoimmunity and through the progression of autoimmunity to hyperglycemia and clinical manifestations. The variations in microbiome composition and dysbiosis, defined by decreased microbial diversity and the relative adequacy of SCFAs-producing anti-inflammatory bacteria, were revealed before and after the onset of islet autoimmunity. Microbiome metabolites synthesized upon the degradation of dietary fibre and starch, such as the SCFAs butyrate, propionate, and acetate, play pivotal roles in gut epithelial integrity and immune homeostasis. However, dysbiosis causes a depletion of SCFAs and decreases epithelial integrity, which paves the way for the mainstream use of dietary antigens and immune stimulants (
Figure 2), thereby aggravating autoimmune diseases such as T1D
| [18] | Harbison JE, Roth‐Schulze AJ, Giles LC, Tran CD, Ngui KM, Penno MA, et al. Gut microbiome dysbiosis and increased intestinal permeability in children with islet autoimmunity and type 1 diabetes: A prospective cohort study. Pediatr Diabetes 2019: pedi.12865. https://doi.org/10.1111/pedi.12865 |
[18]
.
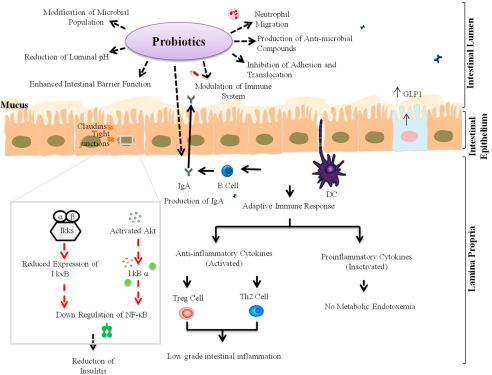
Fortunately, probiotics, to the other extreme, are a group of microorganisms and vital components of the gut microbiota that play critical roles in modulating healthy gut microbiota homeostasis and the production of essential metabolites. Probiotics can exert their effects in many ways, such as by controlling the composition and activation of the metabolic profile of the intestinal microbiota, ameliorating intestinal barrier function and integrity, and stimulating molecules that influence humoral and cellular immune responses
| [15] | Dovi KS, Bajinka O, Conteh I. Evidence and possible mechanisms of probiotics in the management of type 1 diabetes mellitus. J Diabetes Metab Disord 2022; 21: 1081–94. https://doi.org/10.1007/s40200-022-01006-2 |
[15]
. A number of studies, both in vivo and in vitro, have demonstrated the efficacy of probiotics in modulating intestinal epithelial composition, integrity and treatment and/or protection against T1D.
2.1.1. Animal Model Studies
The mode of action of probiotics in T1D treatment is either reducing blood glucose levels or enhancing insulin secretion to maintain the required level. Moreover, ensuring epithelial mucosal membrane integrity through an exogenous probiotic supply is a major intervention in T1D disease management
| [12] | Mishra, Wang S, Nagpal R, Miller B, Singh R, Taraphder S, et al. Probiotics and Prebiotics for the Amelioration of Type 1 Diabetes: Present and Future Perspectives. Microorganisms 2019; 7: 67. https://doi.org/10.3390/microorganisms7030067 |
[12]
. In support of this, it has been possible to modulate intestinal mucosal microbiota diversity and other essential molecules, as demonstrated by the animal model study of
| [19] | Valladares R, Sankar D, Li N, Williams E, Lai K-K, Abdelgeliel AS, et al. Lactobacillus johnsonii N6.2 Mitigates the Development of Type 1 Diabetes in BB-DP Rats. PLoS ONE 2010; 5: e10507. https://doi.org/10.1371/journal.pone.0010507 |
[19]
, in which the administration of
L. johnsonii delayed the onset of T1D in diabetes-resistant rats and markedly altered the intestinal mucosal composition. Blood glucose reduction, as one approach for treating T1D, was possible in an animal study in which probiotic strains of
Lacticaseibacillus paracasei and
Lacticaseibacillus rhamnosus were fed to streptozotocin (STZ)-induced diabetic mice. Along with a decrease in FBG, mice exhibit improvements in body weight, glucose tolerance, insulin levels, and lipid profiles. This study further confirmed that islet integrity of the β-cell mass was enhanced and that access to islets was decreased for macrophages, CD4+ T cells, and CD8+ T cells upon consumption of
Lacticaseibacillus paracasei and
Lacticaseibacillus rhamnosus in diabetic mice. Modulation of the course of T1D disease by probiotic strains of Bifidobacteriaceae, Lactobacillaceae, and
Streptococcus thermophilus in a NOD mouse animal model is a promising alternative treatment for T1D
| [20] | Dolpady J, Sorini C, Di Pietro C, Cosorich I, Ferrarese R, Saita D, et al. Oral Probiotic VSL#3 Prevents Autoimmune Diabetes by Modulating Microbiota and Promoting Indoleamine 2,3-Dioxygenase-Enriched Tolerogenic Intestinal Environment. Journal of Diabetes Research 2016; 2016: 1–12. https://doi.org/10.1155/2016/7569431 |
[20]
.
2.1.2. Clinical Trial Studies
In humans, T1D is triggered by both genetic and environmental factors, including complex genetic elements, patient exposure and the gut microbiome. Viral infections and widespread gut dysbiosis are among the frequent causes of T1D
. Metagenomic analysis of the human gut microbiota revealed that the onset of T1D starts during childhood. A stool sample metagenome study indicated the presence of genes responsible for short chain fatty acids. The infant gut is first colonized mostly by Bifidobacterium species (
B. bifidum,
B. breve or
B. longum) or by the phylum Proteobacteria. Therefore, the development and progression of T1D largely depends on, as confirmed by animal model studies, the extent of early childhood colonization and the production of SCFAs by microbes such as Bifidobacterium species
| [22] | Vatanen T, Franzosa EA, Schwager R, Tripathi S, Arthur TD, Vehik K, et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature 2018; 562: 589–94. https://doi.org/10.1038/s41586-018-0620-2 |
[22]
. The risk of T1D in the infant gut microbiome can be improved by feeding probiotics. Early supplementation with probiotics meaningfully reduced islet autoimmunity in infants compared with that in infants not fed probiotics
. Similarly, the daily application of capsules containing 10
8 CFU of
L. johnsonii for 8 weeks resulted in an increase in the Th1 cell (CD45RO+CD183+CD196−) and cytotoxic CD8+ T cell subsets and an overall positive impact on the innate and adaptive immune systems, which implies that the consumption of
L. johnsonii probiotics can treat T1D
| [24] | Marcial GE, Ford AL, Haller MJ, Gezan SA, Harrison NA, Cai D, et al. Lactobacillus johnsonii N6.2 Modulates the Host Immune Responses: A Double-Blind, Randomized Trial in Healthy Adults. Front Immunol 2017; 8: 655. https://doi.org/10.3389/fimmu.2017.00655 |
[24]
. Compared with that of the placebo, the synergistic effect of probiotic strains resulted in an optimum reduction in fasting blood glucose. A combination of
Bifidobacterium longum,
Lactobacterium bulagricumi, and
Streptococcus thermophilus strains strongly decreased fasting and postprandial glucose and lipid profiles in adult T1D patients
| [16] | Zhang X, Zhang Y, Luo L, Le Y, Li Y, Yuan F, et al. The Beneficial Effects of a Multispecies Probiotic Supplement on Glycaemic Control and Metabolic Profile in Adults with Type 1 Diabetes: A Randomised, Double-Blinded, Placebo-Controlled Pilot-Study. DMSO 2023; Volume 16: 829–40. https://doi.org/10.2147/DMSO.S400119 |
[16]
.
2.2. Rheumatic Arthritis
Rheumatic arthritis is an autoimmune disease characterized by the production of autoantibodies leading to long-term inflammation of multiple joints. The progressive development of RA results in severe deterioration of cartilage and bone, and the destruction of joint structure, ultimately leading to functional disability. If left untreated, the disease can be complicated by the attack of important internal organs, such as the lungs, heart, and kidneys
| [25] | Ferro M, Charneca S, Dourado E, Guerreiro CS, Fonseca JE. Probiotic Supplementation for Rheumatoid Arthritis: A Promising Adjuvant Therapy in the Gut Microbiome Era. Front Pharmacol 2021; 12: 711788. https://doi.org/10.3389/fphar.2021.711788 |
| [26] | De Oliveira GLV, Leite AZ, Higuchi BS, Gonzaga MI, Mariano VS. Intestinal dysbiosis and probiotic applications in autoimmune diseases. Immunology 2017; 152: 1–12. https://doi.org/10.1111/imm.12765 |
[25, 26]
. Both genetic and environmental factors trigger the development of RA, in which the interaction of HLA genes is aggravated by smoking and infection. In addition, dysbiosis has been pinpointed as a primary factor for initiating autoimmunity and the onset of RA. This disease is the most common ADs and is common among women, with a prevalence of 0.3-1% worldwide
| [27] | Paul AK, Paul A, Jahan R, Jannat K, Bondhon TA, Hasan A, et al. Probiotics and Amelioration of Rheumatoid Arthritis: Significant Roles of Lactobacillus casei and Lactobacillus acidophilus. Microorganisms 2021; 9: 1070. https://doi.org/10.3390/microorganisms9051070 |
[27]
.
The RA inflammatory process is a sum effect of autoantibodies and signal transduction pathways of the innate and adaptive immune system. Fluid accumulation and rapid joint cell differentiation lead to the formation of immune cells that express IL-1, IL-17 and TNF-α. However, imbalances between mediators such as IL-11, IL-13 and IL-10 induce immune responses. The penetration of joints by important immune cells, viz. T cells, B cells, and macrophages result in the formation of proinflammatory cytokines that mediate inflammation and cause tissue damage. TLRs, which are responsible for the recognition of pathogen virulence factors, in joint fluids initiate inflammation that results in characteristic features of RA: swollen joints, discomfort, rigidity, and bone and cartilage deterioration
| [25] | Ferro M, Charneca S, Dourado E, Guerreiro CS, Fonseca JE. Probiotic Supplementation for Rheumatoid Arthritis: A Promising Adjuvant Therapy in the Gut Microbiome Era. Front Pharmacol 2021; 12: 711788. https://doi.org/10.3389/fphar.2021.711788 |
| [28] | Bungau SG, Behl T, Singh A, Sehgal A, Singh S, Chigurupati S, et al. Targeting Probiotics in Rheumatoid Arthritis. Nutrients 2021; 13: 3376. https://doi.org/10.3390/nu13103376 |
[25, 28]
.
Gut microbiota dysbiosis has been identified as the hub of RA disease progression and complications. The immune system is directly influenced by the integrity of the gut microbiota. Reports in scientific publications indicate promising results in the treatment and prevention of RA through the modulation of the gut microbiota
| [29] | Asoudeh F, Djafarian K, Akhalghi M, Mahmoudi M, Jamshidi AR, Farhadi E, et al. The effect of probiotic cheese consumption on inflammatory and anti-inflammatory markers, disease severity, and symptoms in patients with rheumatoid arthritis: study protocol for a randomized, double-blind, placebo-controlled trial. Trials 2022; 23: 180. https://doi.org/10.1186/s13063-022-06113-2 |
[29]
. Patients with RA have been proven to have an imbalanced gut microbiota, which strongly contributes to the onset and progression of the disease, as confirmed by faecal biota analysis of healthy and RA-positive individuals
| [30] | Hatakka K, Martio J, Korpela M, Herranen M, Poussa T, Laasanen T, et al. Effects of probiotic therapy on the activity and activation of mild rheumatoid arthritis – a pilot study. Scandinavian Journal of Rheumatology 2003; 32: 211–5. https://doi.org/10.1080/03009740310003695 |
[30]
. In addition, RA patients exhibit a dramatic decrease in the abundance of Bifidobacterium species and LAB. A dietary supply of probiotics is thus indispensable for proper functioning and composition of the microbiota
| [31] | Abhari K, Shekarforoush SS, Hosseinzadeh S, Nazifi S, Sajedianfard J, Eskandari MH. The effects of orally administered Bacillus coagulans and inulin on prevention and progression of rheumatoid arthritis in rats. Food & Nutrition Research 2016; 60: 30876. https://doi.org/10.3402/fnr.v60.30876 |
[31]
. The consumption of probiotic microorganisms, as confirmed by numerous research reports, plays a crucial role in modulating the gut microbiota in many ways: maintaining the equilibrium between beneficial and harmful microbes and eliminating bacteria responsible for allergies and illnesses
| [28] | Bungau SG, Behl T, Singh A, Sehgal A, Singh S, Chigurupati S, et al. Targeting Probiotics in Rheumatoid Arthritis. Nutrients 2021; 13: 3376. https://doi.org/10.3390/nu13103376 |
[28]
.
The therapeutic effect of a probiotic diet is attributed to the downregulation or upregulation of the immune system, which contributes greatly to eliminating symptoms of RA. Preventing gut permeability through the action of probiotics initiates the production of IgA to hinder the proliferation of pathogens, and suppressing important inflammatory immune factors, such as IFN-g, IL-12, and TNF-a, without altering regulatory cytokines, such as IL-10 and TGF-b, is among the mechanisms of probiotics action
| [31] | Abhari K, Shekarforoush SS, Hosseinzadeh S, Nazifi S, Sajedianfard J, Eskandari MH. The effects of orally administered Bacillus coagulans and inulin on prevention and progression of rheumatoid arthritis in rats. Food & Nutrition Research 2016; 60: 30876. https://doi.org/10.3402/fnr.v60.30876 |
[31]
. The efficacy of probiotics in suppressing inflammation relies on the type of strain. Some strains have been found to hinder immune responses, which is thought to be beneficial for patients who experience disease involving immune dysfunction, including RA
| [32] | Vaghef-Mehrabany E, Alipour B, Homayouni-Rad A, Sharif S-K, Asghari-Jafarabadi M, Zavvari S. Probiotic supplementation improves inflammatory status in patients with rheumatoid arthritis. Nutrition 2014; 30: 430–5. https://doi.org/10.1016/j.nut.2013.09.007 |
| [33] | Mohammed, Khattab, Ahmed, Turk, Sakr, M. Khalil A, et al. The therapeutic effect of probiotics on rheumatoid arthritis: a systematic review and meta-analysis of randomized control trials. Clin Rheumatol 2017; 36: 2697–707. https://doi.org/10.1007/s10067-017-3814-3 |
[32, 33]
.
2.2.1. Animal Model Studies
The therapeutic value of probiotics has been widely studied and documented in numerous scientific studies. Animal model studies and clinical trials have proven promising results in the prevention and treatment of RA. The evaluation of the prophylactic effects of common probiotic species of Bifidobacterium by
| [34] | Achi SC, Talahalli RR, Halami PM. Prophylactic effects of probiotic Bifidobacterium spp. in the resolution of inflammation in arthritic rats. Appl Microbiol Biotechnol 2019; 103: 6287–96. https://doi.org/10.1007/s00253-019-09864-2 |
[34]
through oral administration to a rat model of arthritis revealed a significant reduction in arthritis markers and substantial improvements in bone mineral content, cytokine, and eicosanoid levels. The immunoregulatory impact of
Lactobacillus casei and
Lactobacillus acidophilus was assessed in a collagen-induced arthritis (CIA) experimental model, and
Lactobacillus casei and
Lactobacillus acidophilus were found to downregulate proinflammatory cytokines and upregulate anti-inflammatory cytokines
| [35] | Amdekar, Singh, Kumar, Sharma, Singh. Lactobacillus casei and Lactobacillus acidophilus Regulate Inflammatory Pathway and Improve Antioxidant Status in Collagen-Induced Arthritic Rats. Journal of Interferon & Cytokine Research 2013; 33: 1–8. https://doi.org/10.1089/jir.2012.0034 |
[35]
. The outcome of probiotic therapy largely relys on the progressive stage of RA at which probiotics are employed. The study by
| [36] | Fan, Yang, Ross, Stanton, Shi, Zhao J, et al. Protective effects of Bifidobacterium adolescentis on collagen-induced arthritis in rats depend on timing of administration. Food Funct 2020; 11: 4499–511. https://doi.org/10.1039/D0FO00077A |
[36]
reported that the administration of
Bfidobacterium adolescentis to CIA model rats can hinder the onset of arthritic inflammation, and GM-modulating therapies are effective when applied at an early stage of the disease or even before disease occurrence.
2.2.2. Clinical Trial Studies
RA patients demonstrated a decrease in IL10, IL-12 and TNF-α after taking capsules containing 10
8 CFU of
L. casei for 8 weeks
| [37] | Alipour B, Homayouni‐Rad A, Vaghef‐Mehrabany E, Sharif SK, Vaghef‐Mehrabany L, Asghari‐Jafarabadi M, et al. Effects of L actobacillus casei supplementation on disease activity and inflammatory cytokines in rheumatoid arthritis patients: a randomized double–blind clinical trial. Int J of Rheum Dis 2014; 17: 519–27. https://doi.org/10.1111/1756-185X.12333 |
[37]
. Although promising results have been reported in the treatment and prevention of RA through the administration of probiotic strains, dose determination and time of administration still require detail research
| [38] | Aqaeinezhad Rudbane SM, Rahmdel S, Abdollahzadeh SM, Zare M, Bazrafshan A, Mazloomi SM. The efficacy of probiotic supplementation in rheumatoid arthritis: a meta-analysis of randomized, controlled trials. Inflammopharmacol 2018; 26: 67–76. https://doi.org/10.1007/s10787-017-0436-y |
[38]
. In contrast, patients treated with
Lactobacillus rhamnosus in a randomized controlled trial (RCT) were given capsules containing 10
9 CFU per day for 12 months, and the overall results indicated that there was no significant reduction in inflammatory parameters
| [25] | Ferro M, Charneca S, Dourado E, Guerreiro CS, Fonseca JE. Probiotic Supplementation for Rheumatoid Arthritis: A Promising Adjuvant Therapy in the Gut Microbiome Era. Front Pharmacol 2021; 12: 711788. https://doi.org/10.3389/fphar.2021.711788 |
[25]
. In some cases, the synergistic effect of probiotics has profoundly affected the progression of RA. In this regard, in a randomized and double-blind placebo-controlled study by
| [39] | Cannarella, Mari, Alcântara, Iryioda, Costa, Oliveira SR, et al. Mixture of probiotics reduces inflammatory biomarkers and improves the oxidative/nitrosative profile in people with rheumatoid arthritis. Nutrition 2021; 89: 111282. https://doi.org/10.1016/j.nut.2021.111282 |
[39]
patients were provided with probiotics daily for 60 days. Substantial reductions in inflammatory biomarkers such as tumour necrosis factor alpha (TNF-α) and interleukin 6 (IL-6) and improvements in the oxidative/nitrosative profiles of patients with RA were observed.
Disease-modifying antirheumatic drugs (DMARDs), which control the development and progression of RA symptoms, are used most often for treating RA. Many studies on DMARDs have shown partial restoration of dysbiosis and strong modulatory effects on the immune system
| [40] | Bodkhe R, Balakrishnan B, Taneja V. The role of microbiome in rheumatoid arthritis treatment. Therapeutic Advances in Musculoskeletal 2019; 11: 1759720X1984463. https://doi.org/10.1177/1759720X19844632 |
[40]
. However, due to severe side effects, gastrointestinal toxicity, and the toxic effects of the combined prescription of hydroxychloroquine and chloroquine on hepatic and renal problems, patients are facing several challenges in which patients become tired of and unable to finish long-lasting medication
| [28] | Bungau SG, Behl T, Singh A, Sehgal A, Singh S, Chigurupati S, et al. Targeting Probiotics in Rheumatoid Arthritis. Nutrients 2021; 13: 3376. https://doi.org/10.3390/nu13103376 |
[28]
. The potential alternative is probiotic medication as implied in clinical trail studies. In this regard, a randomized, double-blind, placebo-controlled clinical trial was conducted by
| [32] | Vaghef-Mehrabany E, Alipour B, Homayouni-Rad A, Sharif S-K, Asghari-Jafarabadi M, Zavvari S. Probiotic supplementation improves inflammatory status in patients with rheumatoid arthritis. Nutrition 2014; 30: 430–5. https://doi.org/10.1016/j.nut.2013.09.007 |
[32]
. The study categorized patients in two groups: one group was given
Lactobacillus casei 01 in which some patients were given a 10
8-colony forming unit (CFU) capsule and the others were provided with a maltodextrin capsule for 8 weeks. A notable decrease in proinflammatory cytokines (tumour necrosis factor-a, interleukin-6, and interleukin-12) suggested that probiotics could be applied as an adjuvant therapy. Moreover, some studies have also demonstrated the enhancement of the anti-inflammatory effect of combinations of probiotics and drugs. The study by
| [41] | Pružinská K, Slovák L, Dráfi F, Poništ S, Juránek I, Chrastina M, et al. Enhanced Anti-Inflammatory Effect of the Combination of Lactiplantibacillus plantarum LS/07 with Methotrexate Compared to Their Monotherapies Studied in Experimental Arthritis. Molecules 2022; 28: 297. https://doi.org/10.3390/molecules28010297 |
[41]
RA therapy with a combination of methotrexate and
Lactiplantibacillus plantarum LS/07 (LB) for the treatment of adjuvant arthritis (AA) in rats has shown to impact all inflammatory parameters in favour of new therapeutic approaches using probiotics and drugs. Although promising animal model results have been reported, it is inevitable that confirmatory intensive clinical trials for dose and remedy time determination should be conducted.
2.3. Graves' Disease (GD)
Graves' disease is an autoimmune disorder caused by the overproduction of thyroid hormone as a result of the stimulation of thyrotropin receptors by thyrotropin receptor-activating antibodies
. GD arises from multiple interactions, including interactions among the HLA region, protein tyrosine phosphatase-22 (PTPN22), cluster of differentiation 40 (CD40), cytotoxic T lymphocyte-associated factor 4 (CTLA4 or CD152), thyrotropin receptor (TSHR), thyroglobulin (Tg), FCRL3 (FC receptor-like-3), and the secretoglobulin 3A2 (SCGB3A2) gene encoding secretory uteroglobin-related protein 1 (UGRP), combined with environmental factors
| [43] | Khan M, S. Lone S, Faiz S, Farooq I, Majid S. Graves’ Disease: Pathophysiology, Genetics and Management. In: Gensure R, editor. Graves’ Disease, IntechOpen; 2021. https://doi.org/10.5772/intechopen.98238 |
[43]
.
The disease is clinically manifested by an outraged and overreactive thyroid gland, a fast heartbeat, Graves’ orbitopathy (GO), pretibial myxedema (PTM), and the development of a goiter. Graves’ orbitopathy (GO) arises from complications of GD and seriously affects livelihoods
. In addition, individuals with GD have symptoms of palpitation, increased appetite, weight loss, irritability and metabolic activity abnormalities. The prevalence of GD is 0.5% globally and is expected to increase in the near future
| [44] | Deng Y, Wang J, Xie G, Zou G, Li S, Zhang J, et al. Correlation between gut microbiota and the development of Graves’ disease: A prospective study. iScience 2023; 26: 107188. https://doi.org/10.1016/j.isci.2023.107188 |
[44]
.
The primary cause of GD is the stimulation of thyroid-stimulating hormone (TSH) receptor (TSHR) on the thyroid gland by thyroid receptor autoantibodies (TRAbs). The interaction of antibodies and receptors ignites the overproduction of thyroid hormone and leads to a condition known as hyperthyroidism
. The main actors involved in GD development are autoantibodies against thyrotropin receptor, which can be detected by the establishment of GD in healthy individuals upon the introduction of thyrotropin receptor antibodies to foetus and healthy recipients. When the thyroid gland is activated to consistently produce thyroid hormone, the exudation of pituitary thyrotropin is diminished
| [43] | Khan M, S. Lone S, Faiz S, Farooq I, Majid S. Graves’ Disease: Pathophysiology, Genetics and Management. In: Gensure R, editor. Graves’ Disease, IntechOpen; 2021. https://doi.org/10.5772/intechopen.98238 |
[43]
.
In GD, intrathyroidal lymphocytic penetration is the main step in the pathogenesis of this disease and initiates the production of antibodies and immune mediators (e.g., cytokines and Fas) by thyroid cells. GD patients have been found to have fewer CD4 lymphocytes in the thyroid than in the blood as a result of the overexpression of Fas in intrathyroidal CD4+ T lymphocytes. The expression of genes, particularly the RNASET2-FGFR1OP-CCR6 region at 6q27 and an intergenic region at the 4p14 locus, including CD40, CTLA-4, thyroglobulin, the TSH receptor, and PTPN22, is responsible for autoimmune thyroid disease. In addition, thyroid-stimulating hormone receptor and major histocompatibility complex class II are thought to be related to thyroid stimulating hormone receptor autoantibodies TRAbs. The pathophysiology of GD is further assisted by the conversion of mononuclear cells to CD34+ fibrocytes, which internally produce cytokines such as interleukin 6 (IL-6) and TNF-alpha in orbital tissues
| [46] | Jacobson EM, Tomer Y. The CD40, CTLA-4, thyroglobulin, TSH receptor, and PTPN22 gene quintet and its contribution to thyroid autoimmunity: Back to the future. Journal of Autoimmunity 2007; 28: 85–98. https://doi.org/10.1016/j.jaut.2007.02.006 |
[46]
.
The gut microbiota plays a direct role in the establishment and maturation of the human immune system. The gut microbiota influences both the innate and adaptive immune systems in the gut and distant organs. The development of gut-associated lymphatic tissue (GALT) is mediated by the microbiota, which is the hub of 70% of the immune system. GALT efficiently manages self-antigens through the control of Toll-like receptors in the intestinal mucosa
| [47] | Knezevic J, Starchl C, Tmava Berisha A, Amrein K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020; 12: 1769. https://doi.org/10.3390/nu12061769 |
[47]
. In addition, the microbiota produces metabolites such as short-chain fatty acids utilized by enterocytes as energy sources that fasten intercellular connections. In contrast, loss of integrity in the microbiota community leads to GD due to the generation of self-antigens by post translational modification of proteins, lipopolysaccharide (LPS)-induced Toll-like receptor 4 activation, the induction of a type 1 (Th1) to type 2 (Th2) T helper cell shift, a reduction in the integrity of intercellular junctions (leaky gut), and the induction of transcriptomic, proteomic, and metabolic changes
. The microbiome composition of GD patients deviates from that of healthy individuals, especially for
Faecalibacterium prausnitzii,
Butyricimonas faecalis,
Bifidobacterium adolescentis and
Akkermansia muciniphila | [49] | Shu Q, Kang C, Li J, Hou Z, Xiong M, Wang X, et al. Effect of probiotics or prebiotics on thyroid function: A meta-analysis of eight randomized controlled trials. PLoS ONE 2024; 19: e0296733. https://doi.org/10.1371/journal.pone.0296733 |
[49]
. Maintaining the gut microbiota by far is, therefore, significant in the prevention and treatment of GD. Accumulating research evidence suggests that the consumption of probiotics has a great role in modulating the microbiota in many ways, as illustrated by various animal model and clinical trial studies.
2.3.1. Animal Model Studies
In vivo animal models of GD are generated by the electroporation of DNA plasmids encoding the human TSHR A subunit or the introduction of an adenovirus expressing the human TSHR A subunit to acquire the phenotypic features of GD
. Numerous research reports have illustrated the correlation between microbiota diversity and the pathogenesis of GD. The female BALB/c mice model was divided into four groups and subjected to various treatments: (i) treatment with the antibiotic vancomycin, (ii) treatment with the probiotic consortium Lab4, (iii) treatment with freeze-dried human faecal material from GO patients and (iv) treatment with deionized water (ddH2O) as a control. Lab4 consists of four lactic acid bacteria, viz.
Lactobacillus acidophilus strains (
Lactobacillus acidophilus CUL60 (NCIMB 30157) and CUL21 (NCIMB 30156)),
Bifidobacterium lactis CUL34 (NCIMB 30172) and
Bifidobacterium bifidum CUL20 (NCIMB 30153) were combined in a capsule containing 10
10 CFU. Compared with other treatments, vancomycin treatment significantly reduced microbiota biodiversity and reduced the severity of GD disease. Although probiotic treatment resulted in increased CD25+ Treg cells in orbital draining lymph nodes, the course of GD was intensified
| [50] | Moshkelgosha S, Verhasselt HL, Masetti G, Covelli D, Biscarini F, Horstmann M, et al. Modulating gut microbiota in a mouse model of Graves’ orbitopathy and its impact on induced disease. Microbiome 2021; 9: 45. https://doi.org/10.1186/s40168-020-00952-4 |
[50]
. In another study, feeding mice
Lactobacillus reuteri improved thyroid function by enhancing free T4, the thyroid mass, and other related physiological conditions. This was supposed to be accompanied by activated interleukin-10 and subsequent T-reg cells
| [47] | Knezevic J, Starchl C, Tmava Berisha A, Amrein K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020; 12: 1769. https://doi.org/10.3390/nu12061769 |
[47]
. It is indispensable that intensive clinical trial research should be performed prior to its application in humans.
2.3.2. Clinical Trial Studies
Many clinical trial studies on probiotic microbes and probiotics combined with drugs have been reported. Patients with GD were provided with probiotic strains in combination to evaluate their therapeutic efficacy against GD in clinical studies by
| [51] | Lin B, Zhao F, Liu Y, Wu X, Feng J, Jin X, et al. Randomized Clinical Trial: Probiotics Alleviated Oral-Gut Microbiota Dysbiosis and Thyroid Hormone Withdrawal-Related Complications in Thyroid Cancer Patients Before Radioiodine Therapy Following Thyroidectomy. Front Endocrinol 2022; 13: 834674. https://doi.org/10.3389/fendo.2022.834674 |
[51]
. The research focused on the restoration of gut dysbiosis through the consumption of probiotic preparations containing
Bifidobacterium infantis,
Lactobacillus acidophilus, Enterococcus faecalis, and Bacillus cereus. The result revealed dramatic improvements in gut and oral microbial diversity. In addition, probiotics led to a lack of energy, constipation, weight gain, and dry mouth and decreased the levels of faecal/serum LPS and plasma lipid indicators in thyroid cancer patients. The fT3 level of patients improved with probiotic intake, which might be a probiotic effect on the deiodination of thyroid hormone or the activity of β-glucuronidase and sulfatase enzymes.
A six-month study conducted by
| [52] | Huo D, Cen C, Chang H, Ou Q, Jiang S, Pan Y, et al. Probiotic Bifidobacterium longum supplied with methimazole improved the thyroid function of Graves’ disease patients through the gut-thyroid axis. Commun Biol 2021; 4: 1046. https://doi.org/10.1038/s42003-021-02587-z |
[52]
on GD patients revealed the profound effect of
Bifidobacterium longum along with methimazole on the intestinal microbiota and its metabolites. Groups of patients who were given only methimazole showed significant improvement in the intestinal microbiome, whereas limited improvement in TRAb was observed. Unlike the sole methimazole treatment group, the GD group supplied with combination of probiotic and methimazole has exhibited continued improvement, ultimately the TRAb restored to the healthy statis quo.
Probiotics cannot directly influence thyroid disease; rather, they can manage microbiota and their metabolites, which affect neurotransmitters, important trace elements in blood, and ultimately enhance thyroid function. In support of this, a randomized, single-blind, controlled clinical trial was conducted by
| [53] | Spaggiari G, Brigante G, De Vincentis S, Cattini U, Roli L, De Santis MC, et al. Probiotics Ingestion Does Not Directly Affect Thyroid Hormonal Parameters in Hypothyroid Patients on Levothyroxine Treatment. Front Endocrinol 2017; 8: 316. https://doi.org/10.3389/fendo.2017.00316 |
[53]
on the effect of a probiotic consortium (VSL#3) on thyroid function in patients simultaneously treated with levothyroxine (LT4). The study confirmed an insignificant difference in TSH and overall thyroid function after two months of oral intake of probiotic products and LT4. Although it has been found that probiotic consortia modulate the intestinal microbiota, the major role of probiotics is increasing the bioavailability of levothyroxine (LT4). In addition, it has been suggested that microbes can affect thyroid hormones by controlling iodine uptake, degradation, and enterohepatic cycling. The synergy of probiotics and their function as trace element (selenium, zinc, and copper) reservoirs can maintain steady hormonal function
| [49] | Shu Q, Kang C, Li J, Hou Z, Xiong M, Wang X, et al. Effect of probiotics or prebiotics on thyroid function: A meta-analysis of eight randomized controlled trials. PLoS ONE 2024; 19: e0296733. https://doi.org/10.1371/journal.pone.0296733 |
[49]
. Thus, probiotics can serve as an adjuvant therapy for thyroid disorders and could provide efficient treatment for both GD and commonly administered drugs.
2.4. Systemic Lupus Erythematosus (SLE)
Systemic lupus erythematosus is an autoimmune disease known to cause damage to the skin, kidneys, lungs, joints, heart and brain. Genetic and environmental factors, viral infection, defective apoptosis and exposure to ultraviolet-B waves are driving agents for SLE. The attachment of autoantibodies to nuclear and cytoplasmic antigens is responsible for the progression of SLE. Dysbiosis has been intensively studied and reported to promote the development of SLE
| [54] | Brusca SB, Abramson SB, Scher JU. Microbiome and mucosal inflammation as extra-articular triggers for rheumatoid arthritis and autoimmunity. Current Opinion in Rheumatology 2014; 26: 101–7. https://doi.org/10.1097/BOR.0000000000000008 |
[54]
.
The pathogenesis of SLE is still under investigation. It has been decided that SLE pathogenesis comprises interconnected and highly interactive genetic and environmental factors. Sex, hormonal imbalance history and the hypothalamo–pituitary–adrenal axis defines susceptibility and clinical manifestations of the disease. The impairment of immune regulatory mechanisms, including apoptotic cells and immune complexes, is thought to be a major factor in the establishment of SLE. Immune system-related damage, excessive antigen, the production of surplus T-cell help, abnormal B-cell suspensions, and the transformation of T helper 1 (Th1) cells to Th2 cells, which cause B-cell hyperactivity and the production of pathogenic autoantibodies against nuclear antigens and immune complexes (ICs), are responsible for the onset of SLE
| [55] | Mok CC, Lau CS. Pathogenesis of systemic lupus erythematosus. J Clin Pathol 2003; 56: 481–90. https://doi.org/10.1136/jcp.56.7.481 |
| [56] | Wang Y, Zhu D, Ortiz-Velez LC, Perry JL, Pennington MW, Hyser JM, et al. A bioengineered probiotic for the oral delivery of a peptide Kv1.3 channel blocker to treat rheumatoid arthritis. Proc Natl Acad Sci USA 2023; 120: e2211977120. https://doi.org/10.1073/pnas.2211977120 |
[55, 56]
. Continuous accumulation of IC in some organs is central to the pathogenesis of SLE which promotes and establishes inflammatory conditions, leading to the appearance of important SLE symptoms
| [57] | Liu Z, Davidson A. Taming lupus—a new understanding of pathogenesis is leading to clinical advances. Nat Med 2012; 18: 871–82. https://doi.org/10.1038/nm.2752 |
[57]
.
The pathogenesis of SLE is aggravated by disproportion of the gut microbiota and dysbiosis. SLE patients have a low Firmicutes/Bacteroidetes ratio. A decrease in firmicutes in mice was implied to occur during the progression of the disease, and young female mice with lupus experienced a marked reduction in Lactobacilli and a low Clostridiaceae abundance
. Consequently, dysbiosis exacerbates the clinical manifestations and complications of SLE, as loss of integrity of the intestinal gut microbiota results in low tolerance to self-antigens. Gut microbiota diversity analysis greatly varies among animal models, implying that host-microbe interactions outweigh the diversity of microbes involved in SLE pathogenesis. For instance, following intestinal integrity damage, important gut pathobionts, specifically;
Enterococcus gallinarum and
Lactobacillus reuteri, can access systemic tissue where they interact with immune cells to trigger the type I IFN pathway and autoantibody production. Some strains, such as
Ruminococcus gnavus can interact with native DNA to induce an anti-double-stranded DNA antibody response
| [59] | Kim J-W, Kwok S-K, Choe J-Y, Park S-H. Recent Advances in Our Understanding of the Link between the Intestinal Microbiota and Systemic Lupus Erythematosus. IJMS 2019; 20: 4871. https://doi.org/10.3390/ijms20194871 |
[59]
. In addition, it has already been proven that gut microbes contribute greatly to the maintenance of immune system homeostasis through the production of SCFs, preservation of the epithelial barrier, and metabolic functions
| [60] | Mirfeizi, Mahmoudi, Faridzadeh. Probiotics as a complementary treatment in systemic lupus erythematosus: A systematic review. Health Science Reports 2023; 6: e1640. https://doi.org/10.1002/hsr2.1640 |
[60]
. Thus, restoring the gut microbiota composition and integrity is vital in the treatment of SLE. Evidence from various research reports implies that the long-term consumption of encapsulated probiotics or the consumption of fermented food supplements is crucial for the rehabilitation of the gut microbiota as a potential alternative for treating SLE
| [61] | Guo X, Yang X, Li Q, Shen X, Zhong H, Yang Y. The Microbiota in Systemic Lupus Erythematosus: An Update on the Potential Function of Probiotics. Front Pharmacol 2021; 12: 759095. https://doi.org/10.3389/fphar.2021.759095 |
[61]
.
The efficacy of treatment strategies for multiple systemic clinical manifestations of SLE is limited. Glucocorticoids (GCs), antimalarial medicines, nonsteroidal anti-inflammatory drugs (NSAIDs), immunosuppressive agents and related biologics for improving the immune system are therapeutic alternatives. Nevertheless, long-term treatment of patients with strong side effects, including gastrointestinal reactions, metabolic disorders and infections, is forcing scientists to look for alternative therapies with less severe or mild side effects
. The growing evidence on the relationship between the gut microbiota and SLE pathogenesis suggests that restoring dysbiosis could be a potential treatment approach. The distinctive feature of SLE is autoantibody secretion against vital nuclear and cytoplasmic contents; antibodies bind to double-stranded DNA (dsDNA), SmRNP, SSA/Ro, and SSB/La
. However, the gut microbiota has been found to contain bacterial species that produce anti-dsDNA antibodies in SLE patients, among which
Aggregatibacter actinomycetemcomitans,
Porphyromonas gingivalis, Treponema denticola, and the commensal
Capnocytophaga ochracea were studied by
| [63] | Bagavant H, Dunkleberger ML, Wolska N, Sroka M, Rasmussen A, Adrianto I, et al. Antibodies to Periodontogenic Bacteria are Associated with Higher Disease Activity in Lupus Patients 2019. ClinExpRheumatol. 2019; 37(1): 106–111. |
[63]
. The important roles of probiotic bacteria and other microbes have been intensively studied using animal models and clinical trials and randomized single- and double-blind trials.
2.4.1. Animal Model Studies
The use of induced mice for the production of autoimmune antibodies for the treatment of SLE disease has been extensively employed to study the amelioration of dysbiosis and the prophylactic or therapeutic effects of probiotic microbes. Pristance-induced female BALB/c mice were divided into nine groups: the prophylactic group and the treatment group. Both groups were given
L. rhamnosus, L. delbrueckii and the consortium starting from day 0 for the prophylactic group and after two months for the treatment group in the study conducted by
| [64] | Khorasani S, Mahmoudi M, Kalantari MR, Lavi Arab F, Esmaeili S, Mardani F, et al. Amelioration of regulatory T cells by Lactobacillus delbrueckii and Lactobacillus rhamnosus in pristane‐induced lupus mice model. Journal Cellular Physiology 2019; 234: 9778–86. https://doi.org/10.1002/jcp.27663 |
[64]
. Considerable reductions in lipogranuloma, antinuclear antibodies (ANAs), and anti-dsDNA antibodies were observed in both the prophylactic and treatment groups; however, the effect was more pronounced in the prophylactic group. Moreover, the T reg and fork head box P3 (Foxp3) levels were significantly increased, whereas the interleukin 6 (IL-6) level was markedly reduced. Overall, these results suggest that probiotic strains play a prominent role in boosting Tregs and reducing inflammatory cytokines, ultimately eliminating the severity of SLE.
A similar study by
demonstrated a reduction in renal pathology as a result of SLE complications after the consumption of Lactobacillus consortia in an MRL/lpr mice model. Dramatic deterioration of the gut microbiota, especially in the Lactobacillus group, in patients with nephritis (MRL/lpr) was observed during SLE development. This study focused on ameliorating Lactobacillus by feeding mice consortia of five Lactobacillus strains
(Lactobacillus oris, Lactobacillus rhamnosus, Lactobacillus reuteri, Lactobacillus johnsonii, and Lactobacillus gasseri). The combination improved the tightness of the gut through mucosal colonization and helped the mice develop an anti-inflammatory environment through reducing IL6 and increasing IL10 in the kidney. The renal Treg-Th17 balance changes to the Treg phenotype, helping to cope with the progression of the disease. Another study
| [66] | Cabana-Puig X, Mu Q, Lu R, Swartwout B, Abdelhamid L, Zhu J, et al. Lactobacillus spp. act in synergy to attenuate splenomegaly and lymphadenopathy in lupus-prone MRL/lpr mice. Front Immunol 2022; 13: 923754. https://doi.org/10.3389/fimmu.2022.923754 |
[66]
on the efficacy of five Lactobacillus spp.
on important symptoms of SLE, namely, splenomegaly and lymphadenopathy, in a lupus-prone MRL/lpr mouse model. Researchers have tried to evaluate the potential of each species individually for treating lupus; however, none of them was as effective as the consortium. The factors involved in splenomegaly and lymphadenopathy substantially varied between the mouse models treated with mixed species. Generally, the synergy of Lactobacillus species implies that the application of numerous probiotic bacteria may ameliorate systemic autoimmunity and benefit patients with lupus.
Probiotic supplementation in an SLE-induced mouse model has been shown to have a pronounced effect when it is administered in combination with commonly prescribed drugs.
| [67] | Kim DS, Park Y, Choi J-W, Park S-H, Cho M-L, Kwok S-K. Lactobacillus acidophilus Supplementation Exerts a Synergistic Effect on Tacrolimus Efficacy by Modulating Th17/Treg Balance in Lupus-Prone Mice via the SIGNR3 Pathway. Front Immunol 2021; 12: 696074. https://doi.org/10.3389/fimmu.2021.696074 |
[67]
illustrates the synergistic effect of
Lactobacillus acidophilus and tacrolimus in a Lupus-Prone mouse SLE model. The mice were orally administered 5 mg/kg Tac and/or 50 mg/kg LA daily for 8 weeks. Tac in combination with LAB significantly improved gut dysbiosis and markedly reduced the serum levels of anti-dsDNA antibodies, and immunoglobulin G2a and renal pathology scores. Combination therapy has also induced T reg cells and reduced T helper 17 (Th17) cells. Probiotic consumption in SLE patients is more curative when it is accompanied by common drug medication.
2.4.2. Clinical Trial Studies
A noteworthy change in the gut microbiome related to SLE pathogenesis, which is thought to be the root cause of SLE development, has been observed in patients with SLE. In support of this finding, a lower ratio of Firmicutes/Bacteroidetes was detected in a cross-sectional study of
| [68] | Hevia A, Milani C, López P, Cuervo A, Arboleya S, Duranti S, et al. Intestinal Dysbiosis Associated with Systemic Lupus Erythematosus. mBio 2014; 5: e01548-14. https://doi.org/10.1128/mBio.01548-14 |
[68]
SLE patients than in healthy individuals. A diversity study of gut dysbiosis revealed that the abundances of
Bacteroidetes, Proteobacteria, and Actinobacteria decreased, as did the abundance of Firmicutes. Moreover, in association with the progression of SLE, nine genera,
Klebsiella, Rhodococcus, Prevotella, Flavonifractor, Eubacterium, Eggerthella, and I
ncertae sedis, have been shown to be abundant, while
Pseudobutyrivibrio and
Dialister have been depleted
. Similarly, the progression of SLE pathogenesis is inversely proportional to the abundance of
Bifidobacterium, Firmicutes, and Lactobacillus and the
Firmicutes/Bacteroidetes ratio, whereas disease progression is directly related to
Ruminococcus gnavus, Streptococcus, Campylobacter, Streptococcus anginosus and
Veillonella .
As implied in animal model studies, the ingestion of probiotics can alter the gut microbiome composition and contribute to immune system homeostasis. By doing so, probiotics can ameliorate the course of SLE. Maintaining the gut microbiome balance via the consumption of probiotics can be a novel therapeutic alternative to prevent SLE symptoms and cardiovascular and renal complications
| [61] | Guo X, Yang X, Li Q, Shen X, Zhong H, Yang Y. The Microbiota in Systemic Lupus Erythematosus: An Update on the Potential Function of Probiotics. Front Pharmacol 2021; 12: 759095. https://doi.org/10.3389/fphar.2021.759095 |
[61]
. To further evaluate the efficacy of probiotics in humans, limited research has been conducted in randomized controlled trials (RCTs). In this regard,
| [71] | Widhani A, Djauzi S, Suyatna FD, Dewi BE. Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial. Cells 2022; 11: 3419. https://doi.org/10.3390/cells11213419 |
[71]
investigated the improvement of the gut microbiota through the consumption of symbiotics to alleviate systemic inflammation in SLE patients in a randomized, double-blind, placebo-controlled trial. The patients were given symbiotic capsules containing 3 × 10
9 CFU of
Lactobacillus helveticus R0052 60%,
Bifidobacterium infantis R0033 20%,
Bifidobacterium bifidum R0071 20%, fructooligosaccharides, and
Saccharum lactis for the placebo group for 60 days on a daily basis.
The symbiotic group showed a marked decrease in IL-6, whereas no significant change was observed in the IL-17 level. The
Firmicutes: Bacteroidetes ratio increased, and improvements in the severity of SLE were examined. Overall, these results indicate the promising therapeutic role of the probiotic consortium upon long-term consumption.
It is obvious that increasing the
Firmicutes: Bacteroidetes ratio improves the gut microbiota balance and the production of SCFAs such as butyrate, which help improve immune homeostasis. Butyrate hinders the translocation of nuclear factor kappa-B to the nucleus, mediating the transcription of genes encoding proinflammatory molecules such as IL-6
| [60] | Mirfeizi, Mahmoudi, Faridzadeh. Probiotics as a complementary treatment in systemic lupus erythematosus: A systematic review. Health Science Reports 2023; 6: e1640. https://doi.org/10.1002/hsr2.1640 |
[60]
, as depicted in
Figure 3.
2.5. Inflammatory Bowel Disease (IBD)
Inflammatory bowel disease refers to a cluster of diseases that arise from long-lasting inflammation in the colon and small intestine, viz. ulcerative colitis and Crohn’s disease. If left untreated, the condition may be complicated and may lead to more serious fatal disease, such as colorectal cancer
| [72] | Zhou J, Li M, Chen Q, Li X, Chen L, Dong Z, et al. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat Commun 2022; 13: 3432. https://doi.org/10.1038/s41467-022-31171-0 |
[72]
. The adaptive immune response is thought to be the prime cause of IBD; however, it was found to be due to innate immunity that creates an imbalance between the beneficial microbiome and the commensal microflora in the human gut, known as dysbiosis
| [73] | Roy S, Dhaneshwar S. Role of prebiotics, probiotics, and synbiotics in management of inflammatory bowel disease: Current perspectives. World J Gastroenterol 2023; 29: 2078–100. https://doi.org/10.3748/wjg.v29.i14.2078 |
[73]
.
Inflammatory bowel disease involves several factors such as genetic predisposition, the gut microbiome, environmental factors and immunological abnormalities. According to genome analysis, 240 genetic risk loci have been identified, which 30 of them are associated with Crohn’s disease and ulcerative colitis. Nucleotide-binding oligomerization domain 2 (NOD2) is among the most thoroughly characterized genes showing mutations in Crohn’s disease, such as the 1007fs, R702W and G908R mutations, resulting in varying degrees of disease severity. The former causes a more severe disease phenotype, while the latter boosts the intense inflammatory cytokine response. NOD2 is also engaged in the recognition of bacterial invasion by binding to muramyl dipeptide (MDP) in the bacterial cell wall, which initiates the production of proinflammatory cytokines, such as IL-12. NOD2-mediated activation of MDP also promotes autophagy in intracellular bacteria. Upon bacterial recognition, NOD2 interacts with ATG16L1, which serves as an autophagy machinery. Mutations in ATG16L1, such as the T300A substitution and deletion of ATG16L1, are directly linked to IBD pathogenesis
| [74] | Guan Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. Journal of Immunology Research 2019; 2019: 1–16. https://doi.org/10.1155/2019/7247238 |
[74]
. Similarly, numerous studies have shown that dysfunction of both adaptive and innate immune pathways is among the driving factors of IBD pathogenesis. The primary defence mechanism, also known as innate immunity, comprises the intestinal mucosal barrier, proteins antagonizing bacteria, the acidic environment in the stomach and innate immune cells such as neutrophils, macrophages, DCs and natural killer T cells, and cytokines and molecules. However, adaptive immunity relies on the presence of pathogens that pass through innate defence mechanisms. Disorganized immune defence in IBD patients is characterized by epithelial dysfunction due to excessive mucus and defective repair, which ultimately initiates inflammation via the normal intestinal flora and allows the passage of immune cells through the lamina propria. Proinflammatory cytokines in the local tissue, including TNF, IL-1
β, IFN-
γ, and cytokines of the IL-23/Th17 pathway, are produced in excess
.
Accumulated research has pinpointed the inevitable role of the gut microbiota in the commencement and progression of IBD. As a major environmental factor in IBD pathogenesis, the imbalance between harmful and commensal dysbiosis has been implicated in metagenomic studies of the faecal microbiota of IBD patients and healthy controls. With this analysis, the bacterial load of IBD patients was found to increase as the disease became more severe in both flamed and non-flamed patients. In addition, the composition of the microbiota is unstable and varies greatly compared to that of healthy individuals. Some groups of bacteria belonging to Lachnospiraceae and Bacteroidetes were depleted, while bacterial species in Proteobacteria increased in number and diversity
. The major influence of the commensal microflora is attributed to protection at the mucosal surface and the systemic prevention of enteric pathogenic infection via the production of metabolites that favour or disfavour the immune system. For example,
Bacteroides fragilis synthesizes polysaccharide A to decrease IL-17 production in the intestine and promote the function of IL-10-producing CD4+ T cells
. IBD is an autoimmune disease that arises due to the loss of a strong bond between the intestinal microbiota and the immune system, where the gut microbiota lead the development of the host immune system and, in turn, the host immune system maintains the structure and function of the gut microbiome
| [79] | Nishida A, Inoue R, Inatomi O, Bamba S, Naito Y, Andoh A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin J Gastroenterol 2018; 11: 1–10. https://doi.org/10.1007/s12328-017-0813-5 |
[79]
. Thus, any alteration in the microbiome has a deleterious impact on the immune system and can cause immune-related diseases such as IBD. Therefore, it is crucial to restore and maintain microbiome composition as a primary treatment strategy in IBD patients.
The counts of enteric normal microflora, particularly Bifidobacteria and Lactobacillus, were substantially different between patients with Crohn’s disease and patients with ulcerative colitis compared to healthy controls. Crohn’s disease patients were found to contain fewer Bifidobacteria but no Lactobacillus. Similarly, remarkable reductions in anaerobic bacteria, anaerobic gram-negative bacteria, and Lactobacillus were observed in ulcerative colitis patients. However, the abundances of aerobic bacteria and Enterobacteriaceae remained unchanged. Moreover, associated with a dramatic reduction in anaerobic bacteria and Lactobacillus, IBD patients lose their anti-inflammatory function
. Research reports conducted using animal models and clinical trials have indicated the possibility of restoring the anti-inflammatory function of deteriorated enteric microbiota upon the consumption of probiotic strains either separately or in combination.
2.5.1. Animal Model Studies
To date, studies have been conducted using animal models to determine the therapeutic efficacy of probiotic strains and have shown incredible results.
| [81] | Chen Y, Jin Y, Stanton C, Ross RP, Wang Z, Zhao J, et al. Dose-response efficacy and mechanisms of orally administered CLA-producing Bifidobacterium breve CCFM683 on DSS-induced colitis in mice. Journal of Functional Foods 2020; 75: 104245. https://doi.org/10.1016/j.jff.2020.104245 |
[81]
reported the efficacy of
Bifidobacterium breve CCFM683 in a dextran sulfate sodium (DSS)-induced colitis mouse model (C57BL/6J mice). The model animals were administered a 0.2 ml suspension of 10
6-10
10 Bifidobacterium breve CCFM683c once a day for 2 weeks. Mice exhibit dramatic improvements in the intestinal barrier via increased production of mucin2, goblet cells and tight junction proteins at doses of 10
10 and 10
9 cfu/day. It has also been possible to restore the gut microbiota and downregulate inflammatory cytokines.
| [82] | Komaki S, Haque A, Miyazaki H, Matsumoto T, Nakamura S. Unexpected effect of probiotics by Lactococcus lactis subsp. lactis against colitis induced by dextran sulfate sodium in mice. Journal of Infection and Chemotherapy 2020; 26: 549–53. https://doi.org/10.1016/j.jiac.2020.01.006 |
[82]
performed a similar study on the effect of the common probiotic strain
Lactococcus lactis subsp. Lactis on colitis induced by dextran sulfate sodium in mice that were given 1 mg, 5 mg, 10 mg, 15 mg, or 20 mg of
L. lactis per day for one week. A high dose (20 mg) reduced intestinal inflammation. Important parameters, such as the epithelial barrier, which reduces gut leakage, for the occurrence of IBD were regulated by the application of
Weissella paramesenteroides to
WpK4 in a C57BL/6J female mouse model that received 10
8 cfu/day for 10 days
| [83] | Sandes, Figueiredo, Pedroso, Sant’Anna, Acurcio L, Abatemarco Junior, et al. Weissella paramesenteroides WpK4 plays an immunobiotic role in gut-brain axis, reducing gut permeability, anxiety-like and depressive-like behaviors in murine models of colitis and chronic stress. Food Research International 2020; 137: 109741. https://doi.org/10.1016/j.foodres.2020.109741 |
[83]
. In the same manner,
Lactobacillus bulgaricus was delivered to a mouse model at 10
9 cfu 3 times per week for 12 weeks and was capable of preventing the inflammatory response and colitis-associated cancer, whereas
Lactobacillus plantarum AR113 at a similar dose was administered to a mouse model for 2 weeks to efficiently mitigate gut dysbiosis
| [84] | Silveira DSC, Veronez LC, Lopes-Júnior LC, Anatriello E, Brunaldi MO, Pereira-da-Silva G. Lactobacillus bulgaricus inhibits colitis-associated cancer via a negative regulation of intestinal inflammation in azoxymethane/dextran sodium sulfate model. WJG 2020; 26: 6782–94. https://doi.org/10.3748/wjg.v26.i43.6782 |
| [85] | Xia, Chen, Wang, Yang, Song X, Xiong, et al. Lactobacillus plantarum AR113 alleviates DSS-induced colitis by regulating the TLR4/MyD88/NF-κB pathway and gut microbiota composition. Journal of Functional Foods 2020; 67: 103854. https://doi.org/10.1016/j.jff.2020.103854 |
[84, 85]
.
However, studies have suggested that the application of multiple strains has a more profound therapeutic impact on IBD treatment than single-strain attempts. Many research reports have shown success in improving IBD progression and recovering from dysbiosis through the use of multiple strains in animal models.
| [86] | Li, Hsu W-F, Chang J-S, Shih C-K. Combination of Lactobacillus acidophilus and Bifidobacterium animalis subsp. lactis Shows a Stronger Anti-Inflammatory Effect than Individual Strains in HT-29 Cells. Nutrients 2019; 11: 969. https://doi.org/10.3390/nu11050969 |
[86]
demonstrated the difference between single-strain and double-strain utilization using
Lactobacillus acidophilus and
Bifidobacterium animalis subsp
. lactis. Both strains were capable of enhancing the expression of important inflammatory-related molecules, including interleukin (IL)-8, NF-κB, MAPK, VCAM-1, COX-2, and TLR2.
L. acidophilus has no effect on intercellular adhesion molecule-1 (ICAM-1) but promotes the inhibitory effect of
Bifidobacterium animalis subsp. lactis. Therefore, their combination has revealed outstanding anti-inflammatory activity. Moreover,
| [87] | Chen Y, Zhang L, Hong G, Huang C, Qian W, Bai T, et al. Probiotic mixtures with aerobic constituent promoted the recovery of multi-barriers in DSS-induced chronic colitis. Life Sciences 2020; 240: 117089. https://doi.org/10.1016/j.lfs.2019.117089. |
[87]
investigated the competence of four (P-qua) and three (P-tri) probiotic strains in which male C57BL/6 mice were treated with a combination of three probiotic strains,
B. infantis, L. acidophilus and
E. faecalis, at 1.5×10
9 CFU and
Bacillus cereus (0.5×10
8 CFU) was added for combination treatment. The synergy of both combinations has resulted in the restoration of the intestinal microflora and the recovery of multiple barriers in the inflamed gut. However, this effect was more pronounced in the quadrable treatment in which an aerobic strain (
Bacillus cereus) was added, which is likely an efficient and promising therapeutic adjuvant for IBD treatment. Similarly, they assessed the therapeutic role of VSL#3, a mixture of 8 probiotic bacteria that control the levels of autoimmune cells and T follicular helper (Tfh) cells. C57BL/6 mice received VSL#3 (3×10
9 live bacteria) daily for 60 days, after which the number of T follicular helper (Tfh) cells was measured. After VSL#3 treatment, the immunoglobulin (IgM, IgG and IgA) concentration and Tfh cell number were markedly reduced. Thus, this study confirmed that VSL#3 can downregulate autoimmunity and can be a possible therapeutic alternative for IBD
| [88] | Liu X, Yu R, Zou K. Probiotic Mixture VSL#3 Alleviates Dextran Sulfate Sodium-induced Colitis in Mice by Downregulating T Follicular Helper Cells. CURR MED SCI 2019; 39: 371–8. https://doi.org/10.1007/s11596-019-2045-z |
[88]
. The underlying mechanism of probiotic IBD treatment was investigated by
| [89] | Zhang, Zhao, Zhu, Ma, Ma, Zhang. Probiotic Mixture Protects Dextran Sulfate Sodium-Induced Colitis by Altering Tight Junction Protein Expressions and Increasing Tregs. Mediators of Inflammation 2018; 2018: 1–11. https://doi.org/10.1155/2018/9416391 |
[89]
administering Bifico of
Bifidobacterium,
Lactobacillus acidophilus, and
Enterococcus) to a mouse model. The mixture ameliorated the expression of tight junction proteins (TJs), upregulated the number of Tregs, and reduced the total CD4
+ T-cell number in both the colon and peripheral blood.
The selection of a probiotic strain needs to be considered since efficient control of the parameters of IBD pathogenesis per sea cannot make a strain a potent candidate for IBD treatment. The safety and related side effects should be taken into account; for example, intestinal damage was observed in a mouse model when treated with Bifico, suggesting that the combination might aggravate disease severity in IBD patients
| [89] | Zhang, Zhao, Zhu, Ma, Ma, Zhang. Probiotic Mixture Protects Dextran Sulfate Sodium-Induced Colitis by Altering Tight Junction Protein Expressions and Increasing Tregs. Mediators of Inflammation 2018; 2018: 1–11. https://doi.org/10.1155/2018/9416391 |
[89]
. Regardless of the history of safe use of probiotic strains, it is vital to evaluate new strains for pathogenicity and virulence, and the situation is more intense in unhealthy individuals such as IBD patients. The study of
| [90] | Pradhan D, Singh R, Tyagi A, H.M. R, Batish VK, Grover S. Assessing the Safety and Efficacy of Lactobacillus plantarum MTCC 5690 and Lactobacillus fermentum MTCC 5689 in Colitis Mouse Model. Probiotics & Antimicro Prot 2019; 11: 910–20. https://doi.org/10.1007/s12602-018-9489-5 |
[90]
on the safety and therapeutic efficacy of the probiotics
Lactobacillus plantarum MTCC 5690 and
Lactobacillus fermentum MTCC 5689 were investigated in a mouse model. Fortunately, none of the isolates had any deleterious impact on animal models as biotherapeutics for the amelioration of inflammatory conditions after establishing their efficacy in human clinical trials.
2.5.2. Clinical Trials Studies
Although the exact role of the gut microbiome in IBD pathogenesis has not been fully elucidated, research has confirmed that the microbiota composition significantly varies among healthy controls and IBD patients. Taken together, these findings suggest that altered microbiota or dysbiosis play a role in the onset and perpetuation of IBD. Microflora analysis of IBD patients revealed the dominance of the enteric microflora and a marked decrease in the abundance of probiotic species, most importantly
Bifidobacterium and
Lactobacillus spp. Therefore, great attention has been given to improving the proportion of probiotics in the intestinal microbiome over the last few decades as a means of treating gut disorders. Since then, oral administration of probiotic preparations to prevent and treat gut-related
| [91] | Selvamani S, Mehta V, Ali El Enshasy H, Thevarajoo S, El Adawi H, Zeini I, et al. Efficacy of Probiotics-Based Interventions as Therapy for Inflammatory Bowel Disease: A Recent Update. Saudi Journal of Biological Sciences 2022; 29: 3546–67. https://doi.org/10.1016/j.sjbs.2022.02.044 |
[91]
discomfort has become a common therapeutic practice. Currently, the consumption of probiotics as a therapeutic alternative can adjust important intestinal microbiota functions, such as improving epithelial barrier function, and/or modifying the response of the mucosal and systemic immune systems in a way that attenuates the intestinal inflammatory response
| [92] | Fedorak R, Demeria D. Probiotic Bacteria in the Prevention and the Treatment of Inflammatory Bowel Disease. Gastroenterology Clinics of North America 2012; 41: 821–42. https://doi.org/10.1016/j.gtc.2012.08.003 |
[92]
. Thus, probiotic strains have been verified for treating IBD in human volunteers at different dosages either in combination or as a single strain alone at different frequencies.
In this regard, a randomized, double-blinded, placebo-controlled efficacy study of
Bifidobacterium longum 536 was performed by
| [93] | Tamaki H, Nakase H, Inoue S, Kawanami C, Itani T, Ohana M, et al. Efficacy of probiotic treatment with Bifidobacterium longum 536 for induction of remission in active ulcerative colitis: A randomized, double-blinded, placebo-controlled multicenter trial. Digestive Endoscopy 2016; 28: 67–74. https://doi.org/10.1111/den.12553 |
[93]
which patients were provided with 2–3 × 10
11 freeze-dried viable cells for 8 weeks. Analysis of the disease activity index (UCDAI) revealed a significant difference between patients administered the probiotic strains and those in the placebo group, with no deleterious side effects. Similarly,
| [94] | Sun Y-Y, Li M, Li Y-Y, Li L-X, Zhai W-Z, Wang P, et al. The effect of Clostridium butyricum on symptoms and fecal microbiota in diarrhea-dominant irritable bowel syndrome: a randomized, double-blind, placebo-controlled trial. Sci Rep 2018; 8: 2964. https://doi.org/10.1038/s41598-018-21241-z |
[94]
determined the potential of
Clostridium butyricum (CB) in treating diarrhea-predominant IBS (IBS-D) when given to patients for four weeks. The strain successfully treated the syndrome as measured by changes in IBS symptoms, quality of life, stool consistency and frequency from baseline. Compared with placebo, CB is more effective at improving overall IBS-D symptoms.
In addition, after prolonged drug treatment, some UC patients can undergo surgery as a part of treatment, which may cause complications known as pochitis, the most common idiopathic inflammation of the ileal pouch mucosa. Scientific speculations suggest that pochitis arises from dysbiosis and related loss of essential molecules for stable immunity. In a study conducted by
where patients were given 2 mg of the probiotic
Clostridium butyricum MIYAIRI (CBM) three times per day for 24 months after hospital discharge. The application of
Clostridium butyricum MIYAIRI (CBM) sufficiently modulates the intestinal microbiota to eliminate the chance of pouchitis. For example, it was possible to reduce the amount of
E. coli that has been reported to propagate in the inflammatory mucosa of patients with UC. Furthermore,
Clostridium butyricum MIYAIRI (CBM) produces SCFAs that are important for boosting immunity and controlling pathogenic microbes in the intestine to reduce the occurrence of pouchitis.
When a single strain formulation of probiotics is not effective in treating and controlling the relapse of IBD, a combination of strains or a combination of probiotics is employed for sufficient therapeutic procedures. The recurrence of Crohn’s disease after surgery was prevented by VSL#3, a mixture of 8 different bacterial probiotic species, viz.
Lactobacillus, 3 strains of
Bifidobacterium, and 1 strain of
Streptococcus salivarius subspecies
thermophiles were used. Although patients who received VSL#3 had reduced levels of mucosal inflammatory cytokines, the differences in Crohn’s disease activity index and inflammatory bowel disease quality of life scores were not significant compared to those in the placebo group
| [95] | Fedorak, Feagan, Hotte, Leddin, Dieleman, Petrunia DM, et al. The Probiotic VSL#3 Has Anti-inflammatory Effects and Could Reduce Endoscopic Recurrence After Surgery for Crohn’s Disease. Clinical Gastroenterology and Hepatology 2015; 13: 928-935. e2. https://doi.org/10.1016/j.cgh.2014.10.031 |
[95]
.
The efficacy of combined probiotic formulations can be enhanced when probiotics are administered in combination with prebiotics for the treatment and prevention of IBD.
| [96] | Kamarli Altun H, Akal Yildiz E, Department of Nutrition and Dietetics, Eastern Mediterranean University School of Health Sciences, Famagusta, North Cyprus, Akin M, Department of Gastroenterology, Akdeniz University School of Medicine, Antalya, Turkey. Effects of synbiotic therapy in mild-to-moderately active ulcerative colitis: A randomized placebo-controlled study. Turk J Gastroenterol 2019; 30: 313–20. https://doi.org/10.5152/tjg.2019.18356 |
[96]
studied the effects of probiotics and prebiotics and reported that probiotics include (3x10
9 CFU)
Enterococcus faecium,
Lactobacillus plantarum,
Streptococcus thermophilus,
Bifidobacterium lactis,
Lactobacillus acidophilus, and
Bifidobacterium long and prebiotics of fructooligosaccharide (225 mg/tablet). The symbiotic agent was delivered to patients with active UC two times daily for 8 weeks. According to the clinical and endoscopic activity of the disease, a considerable number of patients achieved remission compared to patients in the placebo control group. Generally, it was observed that the prescription of a symbiotic along with a common drug has a more profound effect on active UC patients than does independent administration. In the same manner, a multistrain probiotic trial was performed by
| [97] | Pilarczyk Zurek M, Zwolinska Wcisło M, Mach T, Okon K, Adamski P, Heczko PB, et al. Influence of Lactobacillus and Bifidobacterium Combination on the Gut Microbiota, Clinical Course, and Local Gut Inflammation in Patients with Ulcerative Colitis: A Preliminary, Single-center, Open-label Study. J Prob Health 2017; 05. https://doi.org/10.4172/2329-8901.1000163 |
[97]
where a probiotic consortium of
Lactobacillus plantarum,
Lactobacillus rhamnosus, and
Bifidobacterium longum was used
along with commonly prescribed antibiotics/drugs, namely, mesalazine and ciprofloxacin, for 2 months. As a result, the intestinal microbiota showed variation; for example, Lactobacillus species increased, while the abundance of gram-negative species decreased according to the faecal sample analysis.
| [98] | Palumbo VD, Romeo M, Gammazza AM, Carini F, Damiani P, Damiano G, et al. The long-term effects of probiotics in the therapy of ulcerative colitis: A clinical study. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 2016; 160: 372–7. https://doi.org/10.5507/bp.2016.044 |
[98]
also illustrated the effect of probiotics and mesalazine 1200 mg daily for two years. Researchers have confirmed that long-term treatment with anti-inflammatory drugs combined with probiotics is efficient and could be a potential substitute for existing corticosteroid-based medicine for mild-to-moderate ulcerative colitis.